



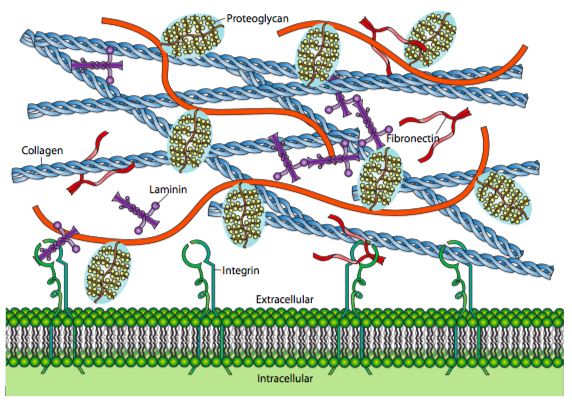
Introducing the Extracellular Matrix
What do you know about extracellular spaces? Do you know that the components of the Extracellular Matrix (ECM) guide tissue morphogenesis, development, and homeostasis, and through the regulation of cellular physiology, growth, survival, differentiation, and adhesion? The ECM is finely tuned to meet local tissue functions influencing chemical and biomechanical signaling for tissue adaptation. Components of the ECM include proteoglycans, glycosaminoglycans (GAG), collagen, and elastin, to name a few. The ECM provides separation between fibrous structures on a macroscale and cell bodies on a micro-scale allowing for potential mobility/slide/glide between structures. Another responsibility for the ECM is providing mechanical resistance to compression along with hydration of the tissues. Alteration of this component is likely the culprit when we feel differences in tissue texture when palpating muscles, fascia, joint capsules, and skin, among others. In particular, the identification of a fluid layer has helped to demonstrate highly organized relationships within the muscular fascia and between the muscle/fascia interface. Before we continue with the ECM, I would like to talk a little about fascia. Fascia? Did somebody say fascia? Please, don’t bore me with that esoteric crap because everyone knows you can’t stretch the iliotibial band.
Fascia and Movement
There has been an increasing number of papers referencing “fascia” on PubMed with movement dysfunction and musculoskeletal pain, and how fascia functions in the body. We generally think about deep fascia from its fibrous components but rarely consider the extracellular components. Since the 30-plus years that I graduated from PT school, not much has changed in the curricula considering the makeup of fascia and its relationship to movement. We still look at the structure grossly as a tissue of containment and separation. PT students still learn that fascia is an inert structure made up of irregular dense connective tissue. Boy, have things changed! It’s time to educate the young and old alike.
 It turns out the muscular fascia is highly organized. Depending on where in the body you look, the muscular fascia is a multilayered structure consisting of 2-3 layers separated by a layer of loose connective tissue. Each fibrous layer contains unorganized parallel collagen fibers in an irregular pattern. Fascia appears to have an irregular pattern because the adjacent layer of parallel collagen fibers has a different orientation/direction. This allows the fascia to give directional resistance in precise patterns. In addition to this microscopic view, we can see that deep fascia macroscopically does more than just contain a muscle, tendon, or joint. It has precise locations where the deep fascia is connected to muscle, where it is connected to the bone, where it is connected to the joint capsule, but just as important, locations where it is free to glide. We have also learned that mechanoreceptor endings, Pacini, Ruffini, and free nerve endings, actually sit within these fascial layers. It is tension through the deep fascia that allows these receptors to fire. In addition, it has been observed that muscle spindles have attachments to the deep fascia and not directly to the extrafusal fibers. Again, tension through the deep fascia allows the muscle spindle to sense a stretch through movement or to adequately contract intrafusal fibers through the gamma motor pathway.
It turns out the muscular fascia is highly organized. Depending on where in the body you look, the muscular fascia is a multilayered structure consisting of 2-3 layers separated by a layer of loose connective tissue. Each fibrous layer contains unorganized parallel collagen fibers in an irregular pattern. Fascia appears to have an irregular pattern because the adjacent layer of parallel collagen fibers has a different orientation/direction. This allows the fascia to give directional resistance in precise patterns. In addition to this microscopic view, we can see that deep fascia macroscopically does more than just contain a muscle, tendon, or joint. It has precise locations where the deep fascia is connected to muscle, where it is connected to the bone, where it is connected to the joint capsule, but just as important, locations where it is free to glide. We have also learned that mechanoreceptor endings, Pacini, Ruffini, and free nerve endings, actually sit within these fascial layers. It is tension through the deep fascia that allows these receptors to fire. In addition, it has been observed that muscle spindles have attachments to the deep fascia and not directly to the extrafusal fibers. Again, tension through the deep fascia allows the muscle spindle to sense a stretch through movement or to adequately contract intrafusal fibers through the gamma motor pathway.
Fascia and ECM
Ok, what does this loose connective tissue layer, fascia, and ECM have to do with each other? Well, it turns out individual cells are reasonably adept at sensing the environment in which they reside. On a microscopic level, the changes in stiffness of the ECM can influence cell migration, cell proliferation, or even apoptosis or cell death. Proteoglycans are rapidly emerging as dynamic modulators of normal states (i.e., ECM hydration, supramolecular assembly, homeostasis, development, wound healing, tissue repair, and senescence) and pathobiological conditions (i.e., inflammation, autophagy, fibrosis, osteoarthritis, atherosclerosis, and cancer). From a macroscopic perspective, changes in the viscosity of the loose connective tissue influence the viscoelasticity, or slide/glide, of the deep fascia layers on each other along with the fascia’s ability to slide over a muscle. Macroscopically we can also see how it impacts the ability of muscle fibers to slide on each other and, on an even smaller scale, impact the contraction of the muscle spindle.
Examining the GAG Hyaluronan
The GAG hyaluronan (HA) is of particular interest regarding changes in viscoelasticity. It contributes to water retention in tissues and their structural integrity and plays a pivotal role in embryogenesis and tissue repair, regeneration, and homeostasis. Hyaluron has versatile functions depending on its size, concentration, and interaction with cell receptors and its ECM binding partners. One HA molecule of high molecular weight HMWHA binds 25 molecules of water! It is hydrophilic or “water-loving” and helps to maintain a specific tissue viscosity or an anticipated slide/glide.
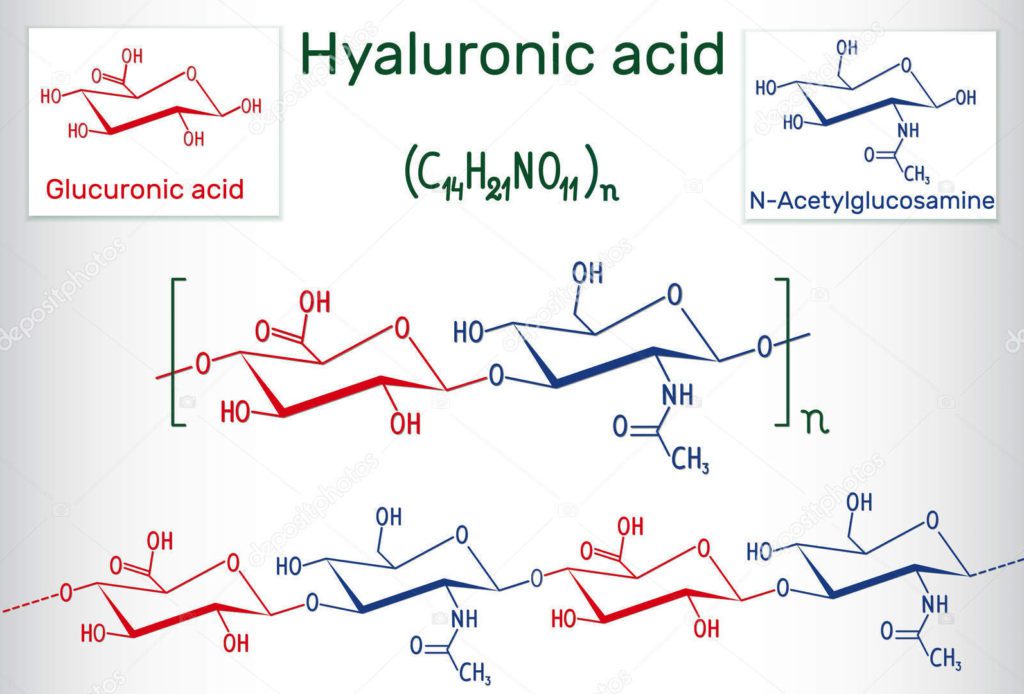 Hyaluronan is susceptible to physical and chemical stressors, such as lactic acid, which can impact its molecular weight and hence its ability to bind water. When this happens, viscosity is altered, and tissue “stiffness” increases, affecting its ability to slide/glide. Likely, we have all experienced soreness following an intense workout or activity and assumed the soreness was due to lactic acid production; however, there is research that suggests that the actual stiffness/soreness we experience may be due to changes in tissue viscoelasticity as a result of a drop of HA molecular weight. This can be due to chemical stressors, such as increased lactic acid production and hence a lowered pH in the local tissues. Lactic acid evacuation happens rather quickly; however, there is a 2-3 day process for HA to turnover and restore local ECM homeostasis. This timeline mirrors the “muscle soreness” most folks experience after intense activity.
Hyaluronan is susceptible to physical and chemical stressors, such as lactic acid, which can impact its molecular weight and hence its ability to bind water. When this happens, viscosity is altered, and tissue “stiffness” increases, affecting its ability to slide/glide. Likely, we have all experienced soreness following an intense workout or activity and assumed the soreness was due to lactic acid production; however, there is research that suggests that the actual stiffness/soreness we experience may be due to changes in tissue viscoelasticity as a result of a drop of HA molecular weight. This can be due to chemical stressors, such as increased lactic acid production and hence a lowered pH in the local tissues. Lactic acid evacuation happens rather quickly; however, there is a 2-3 day process for HA to turnover and restore local ECM homeostasis. This timeline mirrors the “muscle soreness” most folks experience after intense activity.
The Source of Pain and Stiffness
Why does this change in ECM viscosity produce pain or a sense of stiffness? Let’s revisit the fact that our mechanoreceptors actually sit within the deep fascia. If the ability of the deep fascia and muscle to slide/glide is typically impacted, there will be a change in the normal stimulation of these receptors. The brain, through cortical mapping, has an expectation which mechanoreceptors will be firing with planned movement. When the threshold for firing is altered due to changes in tissue stiffness, the brain labels the altered afferents. Sometimes the brain labels the sensation as a feeling of being “stiff” or “tight” as we try to move. Sometimes the abnormal afferents are perceived as pain.
Interestingly, imaging via diagnostic ultrasound (US) and now also using T1 (T rho) MRI show alterations in this ECM component and, more specifically, in HA. Studies have shown changes in echogenicity in muscles for many years when studying myofascial trigger points (TrPs) via US. It appears that alterations in the viscosity of the ECM/LCT have a similar appearance when observed through US. It is widely known that TrPs feature abnormal motor endplate activity and an increase in pro-inflammatory mediators in their immediate milieu. The literature also shows that the HA fragments can be recognized by specific receptors triggering specific inflammatory pathways. Maybe we are talking about similar things when discussing alterations in the ECM and musculoskeletal dysfunction related to TrPs.
If we take time to consider the architecture of the deep fascia, the fibrous component in conjunction with the loose connective tissue, and the ECM, I believe we can have a clearer understanding of which elements are associated with musculoskeletal dysfunction. In future blogs, we will take a closer look at the arrangements of the fibrous components of deep fascia through myotendinous expansions, the relationship between muscle spindles and Golgi tendon organs, and also look at dermatomes, myotomes, and a relatively new concept of fasciatomes.
I would encourage you to investigate the Eureka Seminars’ course offerings in Fascial Manipulation-Stecco Method and take a deep dive into a biomechanical model used for the assessment and treatment of these related structures.
Further Reading
Menon RG, Oswald SF, Raghavan P, Regatte RR, Stecco A. T-mapping for musculoskeletal pain diagnosis: case series of variation of water-bound glycosaminoglycans quantification before and after fascial manipulation® in subjects with elbow pain. Int J Environ Res Public Health. 2020 Jan 22;17(3):708.
Stecco A, Pirri C, Caro R, Raghavan P. Stiffness and echogenicity: Development of a stiffness-echogenicity matrix for clinical problem-solving. Eur J Transl Myol. 2019 Sep 12;29(3):8476.
Cowman MK, Schmidt TA, Raghavan P and Stecco A. Viscoelastic properties of hyaluronan in physiological conditions. F1000Research 2015, 4:622
Martínez Rodríguez R, Galán del Río F. Mechanistic basis of manual therapy in myofascial injuries. Sonoelastographic evolution control. J Bodyw Mov Ther. 2013 Apr;17(2):221-34.
Stecco A, Gesi M, Stecco C, Stern R. Fascial components of the myofascial pain syndrome. Curr Pain Headache Rep. 2013 Aug;17(8):352.
Fede C, Stecco C, Angelini A, Fan C, Belluzzi E, Pozzuoli A, Ruggieri P, De Caro R. Variations in contents of hyaluronan in the peritumoral micro-environment of human chondrosarcoma. J Orthop Res. 2019 Feb;37(2):503-509.
Karamanos NK, Theocharis AD, Neill T, Iozzo RV. Matrix modeling and remodeling: A biological interplay regulating tissue homeostasis and diseases. Matrix Biol. 2019 Jan;75-76:1-11.
Schaefer L, Tredup C, Gubbiotti MA, Iozzo RV. Proteoglycan neofunctions: regulation of inflammation and autophagy in cancer biology. FEBS J. 2017 Jan;284(1):10-26.
Heldin P, Karousou E, Bernert B, Porsch H, Nishitsuka K, Skandalis SS. Importance of hyaluronan-CD44 interactions in inflammation and tumorigenesis. Connect Tissue Res. 2008;49(3):215-8.
Bollyky PL, Bogdani M, Bollyky JB, Hull RL, Wight TN. The role of hyaluronan and the extracellular matrix in islet inflammation and immune regulation. Curr Diab Rep. 2012 Oct;12(5):471-80.
Cowman MK, Lee HG, Schwertfeger KL, McCarthy JB, Turley EA. The content and size of hyaluronan in biological fluids and tissues. Front Immunol. 2015 Jun 2;6:261.
Fede C, Pirri C, Petrelli L, Guidolin D, Fan C, De Caro R, Stecco C. Sensitivity of the fasciae to the endocannabinoid system: production of hyaluronan-rich vesicles and potential peripheral effects of cannabinoids in fascial tissue. Int J Mol Sci. 2020 Apr 22;21(8):2936.
Fascial Manipulation-Stecco method Level 1 and Level 2 course notes.
{kind=link}